
We’re starting a series of reviews of the lab’s favourite papers. Here’s the first review by Shyam Buddh, featuring the famous LTEE lines. Enjoy!
Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations R E Lenski, M Travisano Proceedings of the National Academy of Sciences Jul 1994, 91 (15) 6808-6814
If we classify evolutionary processes along the axis of time, there are two broad categories that emerge – microevolution and macroevolution. Microevolution involves processes that cause changes in gene frequencies from generation to generation, such as selection, mutation, migration, random genetic drift and recombination. Meanwhile, macroevolution refers to the evolutionary changes that happen over millions of years as typically inferred from the fossil record, such as changes in phenotypes between species. Due to its immense time scale, experimental macroevolution was generally considered to be impossible, and the questions of macroevolution – such as whether the rate of change of phenotypes is constant over many generations of evolution – were left to be answered only via indirect inferences from the fossil record.
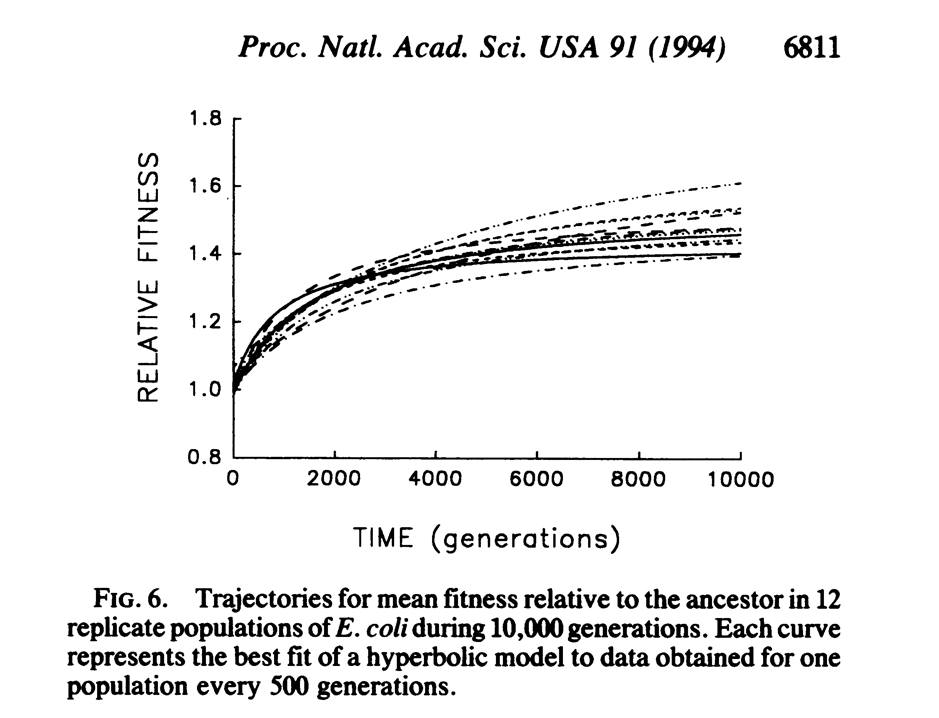
However, in 1994, Richard Lenski and Michael Travisano published a paper in PNAS explaining the results of 10,000 generations of experimental evolution of 12 populations of Escherichia coli on a resource to which they were not adapted. The key thing, as they explained, is that they were able to ‘freeze’ bacterial populations in time, by freezing them at -80 degrees Celsius, and subsequently revive them. This enabled them to examine both the ancestral and derived populations at the same time, and compare the differences in phenotype and fitness. Additionally, they were able to rule out many other factors that affect the conclusions drawn from fossil records – such as potential gene flow between populations, variation in population size over time, and environmental heterogeneity – by carefully controlling the experimental conditions. Hence, using a paleontological analogy, it is as if one has 12 different populations, with identical starting point – all being from the same ancestor – and identical environmental conditions., along with multiple snapshots of their fossils across 10,000 generations. In addition, the fossils can be revived and compared with the ancestors to measure the change in fitness and phenotype over time.
They used this experimental framework to answer many questions of macroevolution, which could not be answered before. Primarily, they asked:
- What are the rates of change in morphology and phenotype, and how do these rates vary over time?
- What is the repeatability of evolution – i.e., how much do populations vary in how they adapt and where they finally reach?
First, they found that the rates of increase of both fitness and morphology (cell size) are very large in the first 2,000 generations, while they are almost zero from generations 5,000 to 10,000. They explained that the reduction in the rate of increase of fitness is likely due to the populations reaching the peak of their adaptive potential, i.e. new mutations no longer cause increases in fitness. Interestingly, even though both fitness and cell size are strongly correlated, they show that it is unlikely that cell size is the primary trait causally affecting fitness, due to the variation in the relationship between cell size and fitness, indicating that it has perhaps actually evolved as a correlated response to selection on some other trait(s).
Secondly, they found that the populations have significantly diverged in their morphology and fitness. In morphology, the populations have continued diverging till generation 10,000, while in fitness, the populations diverged relatively early on and maintained that divergence. The positive association between the rate of increase of fitness and fitness at generation 10,000 suggests that the populations are unlikely to converge in fitness; the authors described this as the populations scaling different peaks on an adaptive landscape.
An important theme in the paper is chance. How much does chance (such as which specific mutations occurred) affect evolutionary trajectories? Given that the different populations reached different peaks while starting as genetically identical clones evolving in the same environment with the same population size, chance is the only factor that can explain the difference in evolutionary trajectories between populations. In fact, the authors showed that every single possible mutation (defined as a change in a single base pair) across the entire genome of the ancestor has probably occurred more than one hundred times over the course of the experiment. This is a tremendous degree of redundancy in mutations between populations. Hence, rather than just which specific mutations were gained, when a population gained the specific mutation is likely to have affected its trajectory and current fitness value – for all populations likely sampled each possible mutation over a hundred times, and hence the more variable factor is when they sampled each mutation, not just the identity of the mutation they sampled.
Two other results from the paper are also quite interesting and worth mentioning. First, they found that in the first 2,000 generations, a step model fits the data well – fitting the classic punctuated equilibrium model of macroevolution. Punctuated equilibrium predicts that macroevolutionary change should happen in periods of rapid bursts that alternate with periods of stasis – and the authors show that the change in fitness over the first 2000 generations fits this model relatively well. Secondly, the variation in the fitness vs time relationship is quite large compared to a smooth, continuously increasing curve – one interpretation of this result is that random genetic drift may play a significant role in determining fitness even with population sizes as high as 500 thousand to 50 million per generation. A second interpretation is that, since the fitness is always measured relative to the 0th generation ancestor, a mutation that increases fitness of one population (the immediate ancestor) could possibly decrease the difference in fitness of that population relative to another population (the 0th generation ancestor).
I think this paper is interesting and significant for taking experimental evolution studies beyond microevolution and into the realm of macroevolution through an original experimental design (which is now used quite commonly, and perhaps could be one of the biggest contributions of this experiment). Additionally, the paper uses concepts from two different scales– macroevolutionary concepts such as chance, history, constraints and necessity, and the adaptive landscape. Using the adaptive landscape as the arena where the macroevolutionary concepts play out creates a link between microevolution and macroevolution; combining that with using macroevolutionary concepts for bacterial evolution really helps connect these two scales of evolution together, and opens up a way to experimentally answer many interesting questions.
