
In the second installment of our series of reviews on the lab’s favourite papers, Shubha Govindarajan writes about a classic paper that gave a new perspective on individual variation and tradeoffs. Read on!
Acquisition and Allocation of Resources: Their Influence on Variation in Life History Tactics by van Noordwijk, Arie & de Jong, Gerdien. (1986), American Naturalist. 128. 137-142
Life history theory attempts to understand the various life history strategies used by an individual (in terms of their reproduction and survival) and its impact on the fitness of the organism. Reproduction and survival- the likelihood of survival of these offspring to adulthood, are considered the most important life history traits for an individual. Since the resources acquired by the individual are distributed between reproduction and survival, a greater resource allocation to reproduction will lead to lower allocation to survival, and vice versa. This is a trade-off- maximizing one trait at the cost of another. While this is the theoretical expectation, there have, surprisingly, been observations of trade-ons between life history traits when trade-offs are expected to be observed.
In the 1970s, work on understanding genotypic and phenotypic correlation among two given life history traits to understand conditions under which trade-offs are likely to arise started gaining importance. However, it was still not very clear as to why trade-offs are, in reality, rarer than they are expected to be.
In 1986, A. J. van Noordwijk, and G. de Jong published a note in The American Naturalist in which they demonstrate using a simple mathematical model, the conditions under which a trade-off between resource allocation to different life history traits is not observed.
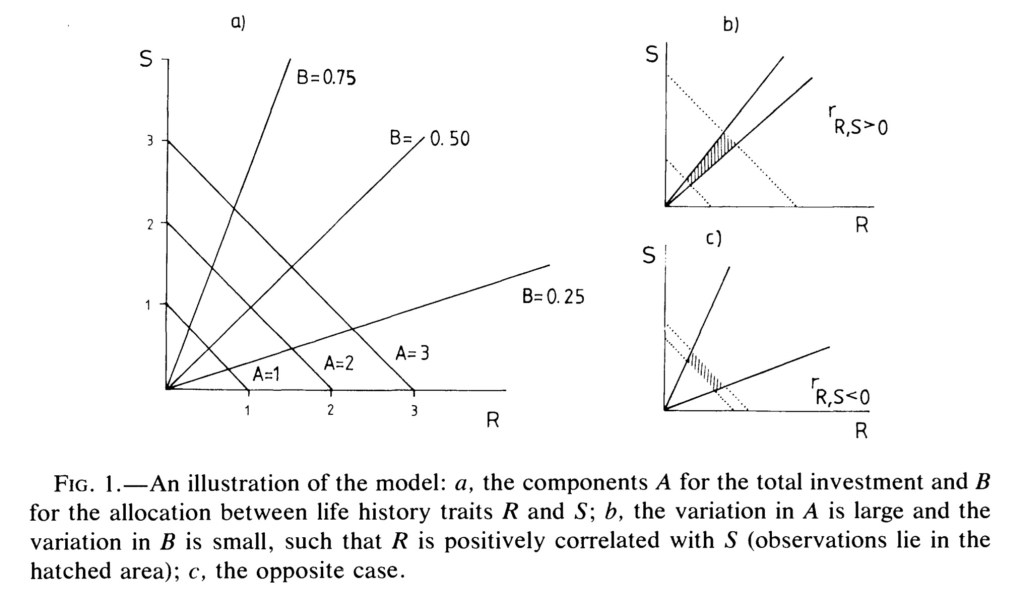
The model developed by the authors describes the total amount of resources available to an individual (A) which is considered as the sum of the amount of resources allocated towards two life history traits, reproduction (R) and survival (S). This relationship between the terms A, R and S (A=R+S) is central to the model, and represents a trade-off between R and S. The correlation between R and S (and consequently, the presence or absence of trade-offs) is given by the measure of covariance between R and S.
(Covariance is a term that describes the sign of the relationship between two variables (that is, whether the variables are directly or inversely proportional to each other). This is different from correlation, which also describes the strength of the relationship of the two variables and is normalized (to the product of the standard deviation of the variables).
If the covariance of A and R is negative, it implies that with increasing energy acquisition, the allocation of energy to reproduction decreases.) This implies that the allocation of energy to survival needs to increase, in order to keep the value of A constant (to satisfy the equation A=R+S) and, hence, a trade-off between R and S. This is more likely to be observed in the long-term, while comparing multiple species or multiple populations, whereas in the short term, at the level of individuals within a population, allocation to reproduction will increase along with the increase in energy acquisition (a trade-on).
But, what about the positive correlations between life history traits when the expectation is to observe negative correlations? The authors consider an aspect of the population that is often overlooked: trait variation among individuals in a population. Individuals in a population may vary in their ability to acquire resources, and also, they may vary in their allocation of resources towards reproduction and survival.
This time, in the model, the authors incorporate a new term- the fraction (B) of acquired energy allocated for reproduction, and the remainder (1-B) allocated for survival. The figure from the paper beautifully illustrates the crux of the model- how the variation in energy acquisition (A) and the fraction of energy allocated to reproduction (B) among individuals in a population play out to influence the covariance between R and S.
- If the variation in A (resource acquisition) is large and that in B (resource allocation) is small, one would observe a positive covariance between R and S, as indicated by the shaded region.
- Conversely, if the variation in A is small and that in B is large, a negative covariance between R and S is observed.
Some individuals in the population may simply be better at acquiring resources. Consequently, if an individual acquires a greater amount of resources, it can produce more offspring which can also survive better (offspring of good quality). This is a positive correlation (and not a trade-off!). So, the authors explain the observation of positive correlations among life history traits by taking into consideration the variation in resource acquisition and allocation among individuals. Hence, one can use the sign of correlation between the allocation of resources to reproduction and survival to deduce the amount of within-individual variation in total energy acquisition and the fraction of allocation towards reproduction (or any life history trait).
The authors use an analogy in economics to explain this observation- given a fixed budget, people who spend more on housing should spend less on cars. However, some families have a greater income, and there is a positive correlation between such families’ expenditures on housing and cars.
In a nutshell, this paper stands out due to its unique way of using a relatively simple mathematical model to explain an observation that is contradictory to what would normally be expected.
(Acknowledgement: I am grateful to Deepa Agashe and Shyamsunder Buddh for discussion on the paper, and for their constructive comments and feedback on the article.)
